
Translate this page into:
Immunoreactivity of glycoproteins isolated from human peripheral nerve and Campylobacter jejuni (O:19)
Address for correspondence: Mrs. Katerina Brezovska, Faculty of Pharmacy, Vodnjanska 17, POB 36, 1000, Skopje, Macedonia. E-mail: kami@ff.ukim.edu.mk
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This article was originally published by Medknow Publications and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Objective:
Antibodies to ganglioside GM1 are associated with Guillain-Barré Syndrome (GBS) in patients with serologic evidence of a preceding infection with Campylobacter jejuni. Molecular mimicry between C. jejuni Lipopolysaccharide (LPS) and ganglioside GM1 has been proven to be the immunopathogenic mechanism of the disease in the axonal variant of GBS. GM1-positive sera cross-react with several Gal-GalNAc-bearing glycoproteins from the human peripheral nerve and C. jejuni (O:19). This study aimed to examine the immunoreactivity of the digested cross-reactive glycoproteins isolated from the human peripheral nerve and C. jejuni (O:19) with Peanut Agglutinin (PNA) as a marker for the Gal-GalNAc determinant, and with sera from patients with GBS.
Materials and Methods:
For this purpose, the cross-reactive glycoproteins from peripheral nerve and C. jejuni (O:19) were enzymatically digested with trypsin and the obtained peptides were incubated with PNA and GBS sera.
Results:
Western blot analysis of the separated peptides revealed several bands showing positive reactivity to PNA and to sera from patients with GBS, present in both digests from peripheral nerve and C. jejuni (O:19).
Conclusions:
These data indicate the possible molecular mimicry between the cross-reactive glycoproteins present in C. jejuni and human peripheral nerve and its potential role in the development of GBS following infection with C. jejuni (O:19).
Keywords
Campylobacter jejuni
Gal-GalNac bearing glycoproteins
Guillain-Barré syndrome
GM1 antibodies
molecular mimicry

Introduction
Guillain-Barré Syndrome (GBS) is the most common cause of acute neuromuscular paralysis. The most frequent variants of GBS include Acute Inflammatory Demyelinating Polyradiculoneuropathy (AIDP) and Acute Motor Axonal Neuropathy (AMAN). About 70% of patients have had a preceding infection. Campylobacter jejuni infection precedes the onset of GBS in 26% of the cases in Western hemisphere[1] and in two-thirds of patients in China, and it seems to be exclusively associated with AMAN.[2] The AMAN variant of GBS is most frequently associated with antibodies to gangliosides GM1, GD1a, and GalNAc-GD1a.[3] The association of GBS with antecedent C. jejuni infection proposed the mechanism of molecular mimicry in the immunopathogenesis of the disease.[4] C. jejuni isolates have been serotyped into about 50 types. A specific serotype of Penner's 19 (O:19) is much more frequently isolated from GBS patients than from enteritis patients.[5] The role of antibodies to the peripheral nerve myelin proteins and glycoproteins was not sufficiently investigated in GBS.[6] GM1-positive sera from patients with GBS following infection with C. jejuni showed reactivity to a 63-kDa flagellar protein purified from C. jejuni (O:19).[7] It has also been shown that GM1 antibodies cross-react with Gal-GalNAc-bearing glycoproteins from the peripheral nerve.[8] We present the results of cross-reactivity of GM1-positive serum with several Gal-GalNAc-bearing glycoproteins isolated from the human peripheral nerve and from C. jejuni (O:19), including the glycoprotein with electrophoretic mobility between 60 and 70 kDa, present in both isolates.[9] These data indicated the possible role of some protein antigens from C. jejuni in the pathogenesis of GBS. Determination of the molecular structure of the glycoprotein structures present in the human peripheral nerve and the bacteria C. jejuni is necessary for elucidation of their antigenicity.
The aim of this study was to examine the reactivity of the peptides obtained after digestion with trypsin of the cross-reactive glycoproteins isolated from the human peripheral nerve and C. jejuni (O:19) with Peanut Agglutinin (PNA) as a marker for the Gal-GalNAc determinant and with sera from patients with GBS.
Materials and Methods
Isolation of glycoproteins from human peripheral nerve and C. jejuni (0:19)
Human peripheral nerve was obtained at autopsy within 8 h after death from patients who died from non-neurological disease; it was kept frozen at -70 °C (Department of Forensic medicine, Faculty of Medicine, University Ss. Cyril and Methodius, Skopje, Macedonia). The neural tissue was pulverized in liquid nitrogen, delipidated with chloroform:methanol (1:2) solution, solubilized by homogenization in 0.5% Triton X-100, 0.4% Sodium Dodecyl Sulfate (SDS) with protease inhibitor cocktail, and heated at 65°C for 10 min. The insoluble matter was removed by centrifugation at 4,200 rpm for 45 min at room temperature.
The C. jejuni serotype (O:19) (obtained from ATCC 43446) was cultured in Campylobacter agar (Campylosel, bioMérieux, France). The bacteria were grown at 37°C for 48 h under microaerophilic conditions. The identity of C. jejuni was confirmed by microscopic examination, by determining the mobility, staining according to Gram and with biochemical tests at the Institute for Microbiology and Parasitology, Faculty of Medicine, Skopje. Bacterial cells were collected in 0.9% NaCl and centrifuged at 4,000 rpm for 30 min. Pellets were resuspended in 8.0 ml of 0.1 M Tris-HCl (pH 7.8) and disrupted by a ultrasonic cell disruptor (MICROSON®, ultrasonic cell disruptor XL, Misonix Incorporated, New York, USA). After centrifugation (45 min, 4,200 rpm), the proteins in the supernatant were dialyzed twice against 0.1M Tris-HCl (pH 7.5) at 4°C for 3 h.[10]
Purification of Gal-GalNAc–bearing glycoproteins
Gal-GalNAc-bearing glycoproteins from the human peripheral nerve and C. jejuni (O:19) were purified by affinity chromatography, using agarose-bound PNA.[8]
Western blot analysis of isolated glycoproteins
Purified glycoproteins isolated from the peripheral nerve and C. jejuni (O:19) were separated on 7.5% acrylamide/bisacrylamide gel (20 μg total glycoproteins per well, 1 μg purified glycoprotein per well) by SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE) and transferred electrophoretically onto nitrocellulose sheets. Unreactive binding sites were blocked in 8% Bovine Serum Albumin (BSA) in Tris-Buffered Saline (TBS - 0.02 M Tris base, 0.5 M NaCl, pH 7.5) for 1 hour at room temperature. The blots were washed three times with TBS containing 1% Tween 20 and incubated overnight at 4°C with biotinylated PNA (Sigma-Aldrich) (10 μg/ml) and with sera from 10 patients with GBS and high titer of anti-GM1 antibodies (Institute of Neurology, Clinical Center of Serbia, Belgrade, Serbia) diluted 1:100. After washing, the strips were incubated with avidin conjugated with peroxidase (1:1000), and with anti-human IgG antibodies conjugated with peroxidase (1:500) for 1 hour at room temperature. Visualization of the reaction products was done using Diaminobenzidine (DAB), 15 mM imidazole, and 0.025% H2O2 in water, (BioRad Laboratories, Hercules, CA, USA).
Purification of cross-reactive glycoproteins by preparative SDS-PAGE
Gal-GalNAc–bearing glycoproteins were electrophoretically separated using 7.5% acrylamide/bisacrylamide gel (approximately 20 μg total glycoproteins/well). After electrophoresis, a strip from the left side of the gel, including molecular weight marker and first line of the protein sample, was cut with a razor and stained with 0.1% Amido Black, 1% acetic acid, and 40% methanol in water. This strip was used as a reference gel strip to localize the bands of glycoproteins with mobility between 60 and 70 kDa. After aligning the reference strip with the unstained gel portion, the bands of the gel that align with the stained protein of interest were cut from the unstained gel and rinsed with distilled water.
Enzyme digestion of the isolated Gal-GalNAc-bearing glycoproteins
Gels containing proteins of interest were equilibrated in 1 ml 0.125 M Tris (pH 6.8) for 10-15 min and added to the wells of a 15% acrylamide/bisacrylamide gel. Enzyme trypsin in buffer was added to the wells and after 10-15 min electrophoresis was run. After the electrophoretic separation, the gel was stained with Silver stain (Silver Stain Plus, BioRad Laboratories, Hercules, CA, USA) for visualization of the peptides.
Determination of the immunoreactivity of the digested glycoproteins
Following electrophoretic separation, obtained peptides were transferred electrophoretically onto nitrocellulose sheets. Unreactive binding sites were blocked in 8% BSA in TBS (0.02 M Tris base, 0.5 M NaCl, pH 7.5) for 1 hour at room temperature. The blots were washed three times with TBS containing 1% Tween 20 and incubated overnight at 4°C with biotinylated PNA (Sigma-Aldrich) (10 μg/ml) and with the sera from patients with GBS (1:100). After washing, the strips were incubated with avidin conjugated with peroxidase (1:1000), and with anti-human IgG antibodies conjugated with peroxidase (1:500) for 1 hour at room temperature. Visualization of the reaction products was done using DAB, 15 mM imidazole, and 0.025% H2O2 in water (BioRad Laboratories, Hercules, CA, USA).
Results
Isolated glycoproteins were tested on Western blot for their reactivity with PNA, as a marker for Gal-GalNAc determinant and with GM1-positive sera from patients with GBS. PNA staining of the separated glycoproteins isolated from the human peripheral nerve and C. jejuni (O:19) indicate the presence of the Gal-GalNAc determinant in several proteins [Figure 1]. Western blot analysis of the reactivity of isolated proteins to sera from patients with GBS, revealed positive reactivity of one main protein band with mobility between 60 and 70 kDa, present in both isolates, with tested sera from patients with GBS [Figure 2]. Figure 2 also shows positive reactivity of bands with molecular weight lower than 60 kDa in both isolates. Further studies are needed to determine whether these bands are degradation products of the main cross-reactive proteins, or are independent proteins that show positive reactivity with sera from patients with GBS.

- Western blot of isolated glycoproteins from human peripheral nerve (Lane 1) and C. jejuni (Lane 2) stained with biotinylated PNA. The arrow indicates the positive reactivity of bands between 60 and 70 kDa.
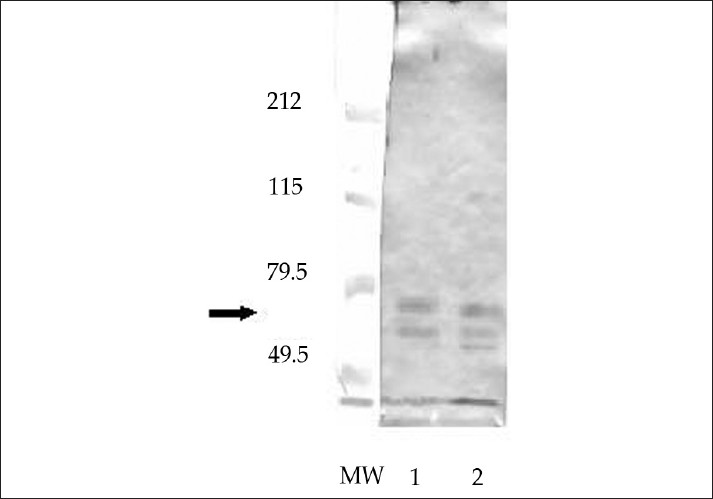
- Western blot of isolated glycoproteins from human peripheral nerve (1) and C. jejuni (2) stained with GM1-positive serum from patient with GBS. The arrow indicates the cross-reactive band between 60 and 70 kDa.
After electrophoretic separation of the glycoproteins isolated from the human peripheral nerve and from C. jejuni (O:19), a reference gel strip cut from the gel was stained with 0.1% Amido Black, 1% acetic acid, and 40% methanol in water [Figure 3]. Bands corresponding to the proteins with mobility of the cross-reactive glycoproteins (between 60 and 70 kDa), as compared to standard, were cut from the unstained gel and used for further enzymatic digestion.
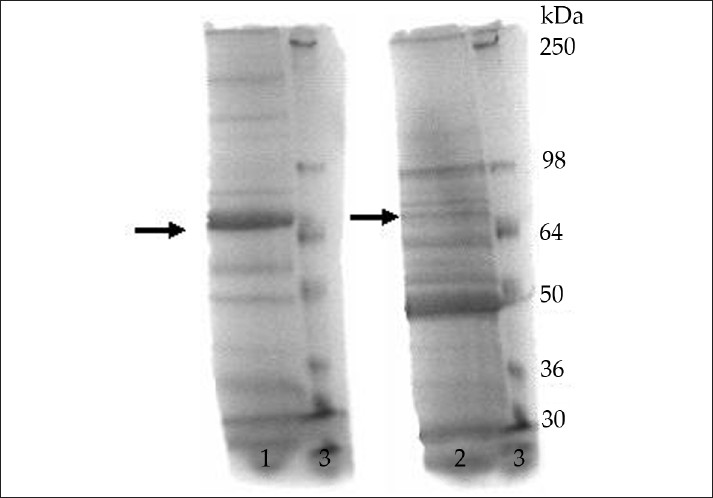
- SDS-PAGE of isolated glycoproteins stained with 0.1% Amido Black, 1% acetic acid, and 40% methanol in water. Lane 1: Human peripheral nerve; Lane 2: C. jejuni (O:19); Lane 3: Protein standard. The arrows indicate the protein bands with mobility between 60 and 70 kDa.
The cross-reactive glycoproteins from human peripheral nerve and C. jejuni (O:19) were enzymatically digested with trypsin and obtained peptides were separated on SDS-PAGE and visualized with Silver stain [Figure 4]. BSA served as a control for digestion and a well without protein served as blank.
- SDS-PAGE of the digested glycoproteins from C. jejuni (Lane 2) and peripheral nerve (Lane 3) stained with Silver stain. Lane 1: BSA; Lane 4: Blank; Lane 5: Protein standard.
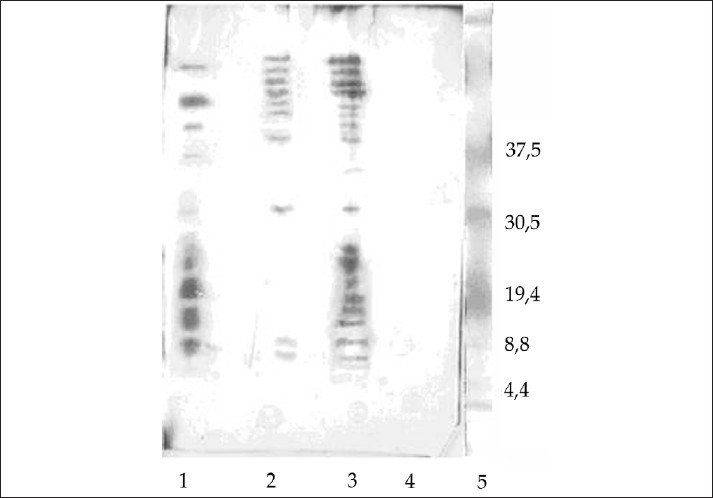
Western blot analysis of the separated peptides obtained after trypsin digestion revealed positive reactivity of several bands to PNA, present in both digests from the peripheral nerve and C. jejuni (O:19), indicating the presence of the Gal-GalNAc residue in these bands [Figure 5]. After incubation with serum from patient with GBS associated with C. jejuni (O:19) infection, bands showing positive reactivity were revealed in both digests [Figure 5]. Serum from healthy individual did not show any reactivity to the obtained peptides (not shown). Further studies are needed to determine the molecular structure of these reactive peptides.

- Western blot of the digested glycoproteins from peripheral nerve (Lane 2) and C. jejuni (O:19) (Lane 3) stained with: (a) GM1 positive serum from patient with GBS, and (b) biotinylated PNA. Lane 1: Peptide standard. The arrow indicates the positively reactive bands.
Discussion
The immunopathogenesis of the AMAN variant of GBS has been very well documented.[11] Infection by C. jejuni PEN 19 that bears a GM1-like Lipopolysaccharide (LPS) induces high production of IgG1 and IgG3 anti-GM1 antibodies. Anti-GM1 antibodies bind to the nodes of Ranvier and motor nerve terminal axons, inhibit motoneuron excitability, and produce muscular weakness. Pathologic studies of early cases of AMAN have shown deposits of activated complement components and immunoglobulins at nodes of Ranvier motor fibers.[12] The disruption of paranodal region allows the entry of complement and immunoglobulins along the axolemma, with subsequent recruitment of macrophages, which invade the periaxonal space, leading to Wallerian degeneration of motor fibers. The immunopathology of GBS associated with C. jejuni infection is caused by a mechanism of molecular mimicry between C. jejuni LPS and GM1 ganglioside of the peripheral nerve. A single C. jejuni strain is shown to have several LPSs with various ganglioside epitopes, and different Penner's serotypes have the same ganglioside epitope.[13] These indicate that Penner's serotyping system is not dependent on ganglioside-like LPSs. On the other hand, some anti-GM1 antibodies may be GM1-monospecific, whereas others may cross-react with other gangliosides or glycoproteins.[14] The subtypes of anti-GM1 antibodies cross-reactive with neural glycoproteins may correlate with different GBS variant.
Our results showed positive cross-reactivity of the peptides from the human peripheral nerve and C. jejuni recognized by GM1-positive GBS serum associated with C. jejuni infection. We have previously shown that GM1 antibodies cross-react with several Gal-GalNAc–bearing glycoproteins at the node of Ranvier in the peripheral nerve. Two of these glycoproteins have been identified as Oligodendrocyte Myelin Glycoprotein (OMGp) localized at the paranodal region and hyaluronate-binding domain of versican localized at the nodal gap.[8] A hyaluronic acid-like repeating unit of LPS is proved to be an antigenic determinant of O: 19.[15] The structural similarity exists between hyaluronic acid-like repeat unit of C. jejuni (O:19) and hyaluronate-binding domain of versican.[8] This supports the hypothesis that pathology in GBS may be mediated by cross-reactive autoantibodies directed against the Gal-GalNAc epitope of ganglioside GM1 and hyaluronate-binding domain of versican, whose production is triggered by homologous antigen in C. jejuni (O:19).
The results of this study showed the presence of cross-reactive determinants in glycoproteins from the human peripheral nerve and C. jejuni (O:19), recognized by PNA, and by GM1-positive serum from patient with GBS, associated with C. jejuni infection. These findings indicate the possible molecular mimicry between isolated cross-reactive glycoproteins and its role in the development of GBS in patients with previous infection with C. jejuni. Further studies are needed to characterize and determine the molecular structure of the obtained cross-reactive glycoproteins and their epitopes.
Conclusions
Elucidation of the molecular structure and function of the cross-reactive glycoproteins present in the human peripheral nerve and C. jejuni will indicate their role in the pathogenesis of GBS and also help in understanding the mechanisms that trigger the immune responses in postinfectious autoimmune diseases. These data indicate the possible molecular mimicry between the cross-reactive glycoproteins present in C. jejuni and human peripheral nerve and also its potential role in the development of GBS following infection with C. jejuni (O:19).
Source of Support: Nil.
Conflict of Interest: None declared.
References
- A prospective case control study to investigate the relationship between Campylobacter jejuni infection and Guillain-Barré syndrome. N Engl J Med. 1995;333:1374-9.
- [Google Scholar]
- Guillain-Barré syndrome in Northern China: Relationship to Campylobacter jejuni infection and anti-glycolipid antibodies. Brain. 1995;118:597-605.
- [Google Scholar]
- Axonal Guillain-Barré syndrome: Relation to anti-ganglioside antibodies and Campylobacter jejuni infection in Japan. Ann Neurol. 2000;48:624-31.
- [Google Scholar]
- Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease. FEMS Immunol Med Microbiol. 1996;16:105-15.
- [Google Scholar]
- Specific serotype of Campylobacter jejuni associated with Guillain-Barré syndrome. J Infect Dis. 1992;165:183.
- [Google Scholar]
- Anti-PMP22 antibodies in patients with inflammatory neuropathy. J Neuroimmunol. 2000;104:139-46.
- [Google Scholar]
- Detection of antibodies against Campylobacter jejuni serogroup PEN O:19 purified flagellar protein in a patient with Guillain-Barré syndrome. Zentralbl Bakteriol. 1999;289:429-44.
- [Google Scholar]
- Identification of Gal (β1-3)GalNAc bearing glycoproteins at the nodes of Ranvier in peripheral nerve. J Neurosci Res. 1994;38:134-41.
- [Google Scholar]
- Motor neuron disease associated with anti-GM1 antibodies crossreactive to the glycoproteins of the human peripheral nerve, spinal cord and Campylobacter jejuni. Belgrade: 35th International Danube Symposium for Neurological Sciences and Continuing Education; Book of Abstracts: . 2003:188-189.
- [Google Scholar]
- Mass Spectrometry-Based Glycomics Strategy for Exploring N-Linked Glycosylation in Eukaryotes and Bacteria. Anal Chem. 2006;78:6081-7.
- [Google Scholar]
- Pathogenesis of axonal Guillain-Barré syndrome: Hypothesis. Muscle Nerve. 1994;17:680-2.
- [Google Scholar]
- Acute motor axonal neuropathy: An antibody-mediated attack on axolemma. Ann Neurol. 1996;40:635-44.
- [Google Scholar]
- Ganglioside-like epitopes of lipopolysaccharides from Campylobacter jejuni (PEN 19) in three isolates from patients with Guillain-Barré syndrome. J Neurol Sci. 1995;130:112-6.
- [Google Scholar]
- Lipopolysaccharides of Campylobacter jejuni serotype O:19: Structures of O antigen chains from the serostrain and two bacterial isolates from patients with Guillain-Barré syndrome. Biochemistry. 1994;33:250-5.
- [Google Scholar]




